Зоогенная сукцессия растительности "сусликовой полупустыни" Северного Прикаспия
Растительный покров полупустыни характеризуется специфической комплексностью и сложным структурным членением фитоценотических группировок (Каменецкая, 1952; Каменецкая, Гордеева и Ларин, 1955; Гордеева, 1959, 1962; Гордеева, Ларин, 1967; Оловянникова, 1966; Левина, 1958, 1960, 1961, 1964; Никитин, 1954, 1960, и др.). Сложность организации растительного покрова определяется как относительным разнообразием самого видового состава растений (значительно превышающим таковое в северных пустынях), так и той мозаичностью условий среды жизни, которую создает специфический мезо- и микрорельеф аллювиальной равнины Северного Прикаспия и во многом соответствующая ему пестрота почв и гидрологических условий (Роде, Польский, 1961).
По данным Ф. И. Левиной (1964), "из 605 видов роль сообществообразователей принадлежит 175 (29,0%) видам 31 семейства (из 67). В это число входит примерно 88 (14,6%) эдификаторов и 87 (14,4%) субэдификаторов основных, наиболее распространенных пустынных, степных, луговых и других растительных сообществ". Видовое разнообразие растений, характеризующееся приведенными цифрами, вместе с тем укладывается в относительно небольшое число основных схем структурной организации растительного покрова, определяющих зональный облик растительности этих глинистых геологически молодых равнин, относительно недавно (около 8 тыс. лет назад) освободившихся из-под вод второй хвалынской трансгрессии Каспийского моря ("плаккаты", по Г. Н. Высоцкому, 1927).
Аквальный характер поверхности легко прослеживается по присутствию в рельефе чередующихся достаточно обширных, но неглубоких западин лиманного типа, что типично для прибрежных зон мелководного морского водоема (Мозесон, 1954, 1956). Однако все особенности рельефа современной поверхности Северного Прикаспия нельзя объяснять ее морским палеогенезисом. В формировании нынешнего мезо- и микрорельефа полупустыни активно участвуют современные биотические (животные и растения) и абиотические факторы. А. А. Роде (1953) указывает на процессы миграции солей в подповерхностном слое солончаковых солонцов, вызывающих вспучивание и поднятие их поверхности. Известное место в динамике рельефа поверхности принадлежит смыву атмосферными осадками и отчасти эоловым процессам.
Весьма существенное значение в формировании современного рельефа имеет активная роющая деятельность грызунов, в первую очередь малых сусликов (Citellus pygmaeus Pall.), создающих холмики выброшенной земли - сусликовины, сохраняющиеся многие десятки и сотни лет (Динесман, 1968; Ходашова, Динесман, 1961; Варшавский, 1960). Суммарная площадь сусликовин - заметных в рельефе холмиков в Джаныбекском районе составляет 4% территории, или 8% площади солончаковых солонцов (Зубкова, 1971) - основного "субстрата", на котором располагаются практически почти все жилые, активно функционирующие сусликовины. Максимальная же суммарная площадь сусликовин в Прикаспии может равняться 5-10% площади (Формозов, Ходашева, Голов, 1954).
Известно, что нарушение поверхности почвы землероями вызывает сложную цепь циклических процессов, изменяет условия водопроницаемости поверхности, регулирует и "организует" сток атмосферной влаги и, следовательно, непосредственно влияет на гидрологический и солевой режим почв (Иозефович, 1928; Иванов, 1950; Ходашова и Динесман, 1961; Абатуров, Зубкова, 1969, 1972; Зубкова, 1971). Растительный покров полупустыни оказывается объектом как прямого влияния грызунов, так и многообразных косвенных воздействий результатов их роющей деятельности.
В настоящее время этим вопросам посвящена уже значительная специальная литература (Воронов, 1954; Формозов, Воронов, 1939; Формозов, Ходашова, Голов, 1954; Варшавский, Ротшильд, 1960; Ротшильд, 1969; Левина, 1964; Залетаев и Зубкова, 1968; Молчанова, 1975, и др.). Однако многие аспекты влияния животных на растительность, на генезис фитоценотических группировок, структуру биогеоценозов и ход сукцессионных процессов, а также биогеографическая оценка зональных схем зоогенных воздействий на растительность аридных земель представляют собой еще обширное поле для исследований.
Исследования воздействия млекопитающих на растительность полупустыни в Северном Прикаспии были начаты автором в 1959 г. и затем после перерыва продолжены в 1967-1974 гг. Наблюдения проводились главным образом в Джаныбекском и Урдинском районах Уральской области Казахской ССР, кроме того, была обследована территория Волжско-Уральского междуречья, от г. Гурьева на север до с. Калмыкове и далее на запад и северо-запад через Фурманово и Александров Гай до г. Энгельса на Волге. Пробные площадки размером от 0,01 га до 0,25 га картировались с инструментальной нивелировкой рельефа, отмечались детали распределения растительности и последствия роющей деятельности малых сусликов (сусликовины, временные норы, прикопки, вертикальные ("колодцы") и наклонные норные ходы, полузаплывшие норы, лункообразные углубления на месте старых, занесенных, обвалившихся и заплывших ходов, главным образом вертикальных, просадки в почве, ваннообразные и нередко подковообразные, образующиеся в случае соединения двух-трех ваннообразных понижений на месте осевшей почвы вокруг норного хода). Особо отмечались в рельефе и растительном покрове следы осевших, давно заброшенных, если так можно выразиться, "руинных сусликовин", возраст которых достигает нескольких сот лет.
Специфическая для жилых сусликовин асимметрия контуров, почвенно-гидрологических показателей и концентрических пятен растительного покрова может быть последовательно прослежена на всех стадиях сложного и длительного цикла модификаций сусликовины, вплоть до момента ее почти полного "поглощения" регенерировавшим покровом чернополынных ассоциаций или в иных случаях при "злаковом поглощении", когда злаки поселяются на образовавшихся круговых, как бы "опоясывающих" сусликовину просадках и затем смещаются на сусликовину, особенно при ее постепенном оседании. В западинах происходит существенное изменение характера увлажнения, а следом и травянистой растительности, которая трансформируется в направлении развития лугово-степных группировок видов. Одновременно начинают формироваться светло-каштановые степные почвы западин.
Стойкость стандартной, типичной только для сусликовин формы асимметрии этих почвенно-растительных концентров позволяет с достаточной степенью уверенности использовать эти чисто морфологические показатели для опознания и выделения мест расположения прежде активных биогеоценотических структур бесспорно зоогенного происхождения.
Развитая сусликовина в возрасте нескольких десятков лет представляет собой холмик выброшенной сусликами и метаморфизировавшейся почвы диаметром 2-4 м и высотой 0,2-0,4 м. В связи с тем что вход в нору располагается обычно не на самой вершине холмика, а несколько смещен к его периферии, выброшенная почва неравномерно распределяется по поверхности первичного бугра, что создает характерную асимметрию. Процесс смыва рыхлого почвенного материала атмосферными осадками не выравнивает поверхность бугра, а нередко усугубляет различия в крутизне склонов относительно односторонне направленным стоком.
Асимметричность формы сусликовин, легко определяемая на глаз, фиксируется при нивелирной съемке местности и продолжается в асимметрии почвенных характеристик различных экспозиций сусликовины, о чем можно судить косвенно по размещению крупных кустов лиственничных солянок (Salsola laricina) и однолетней многолистной солянки (S. fotiosa) и петросимонии (Petrositnonia triandra), поселяющихся на "перезревающих" сусликовинах в возрасте нескольких десятков лет.
Характерный тип асимметрии сусликовин удалось обнаружить при картировании растительности и на участках, где холмики обычной средней высоты отсутствовали, а в микрорельефе возможно было заметить лишь слабо приподнятые пуповиноподобные вздутия поверхности, однако с тем же типичным контуром, повторенным и подчеркнутым топографией растительных группировок. Эти "руинные бутаны" располагались в периферийной зоне злаковых западин, где жилые сусликовины никогда не встречаются из-за весеннего затопления понижений в рельефе. При этом в зависимости от индивидуального возраста почвенно-растительного комплекса, представляющего собой отпечаток прежней деятельности сусликов, состав видов в группировках растений может быть различным, а рисунок структуры остается неизменным. Столь же устойчиво индицируется растительностью и типичная подковообразная форма "опоясывающих" сусликовину (или ее след) просадок, занятых то злаково-ромашниковым покровом, то злаками.
В объективном характере выделенного морфоструктурного признака убеждает отражение его на картосхемах разными исследователями, не ставившими перед собой задачу установить преемственность между молодыми зоогенными структурами растительного покрова и комплексностью полупустыни или вообще не включавшими деятельность животных в сферу своих наблюдений. Так, асимметричная форма сусликовин, их растительного покрова и пятен "послесусликовинной растительности" хорошо видны на схеме заповедного участка полупустыни (Джаныбекский стационар) по съемке в 1952 г. (Роде, Польский, 1961) и на картосхемах, снятых Ф. Я. Левиной (1964) в различных районах полупустыни Волжско-Уральского междуречья (см. рис. 22).
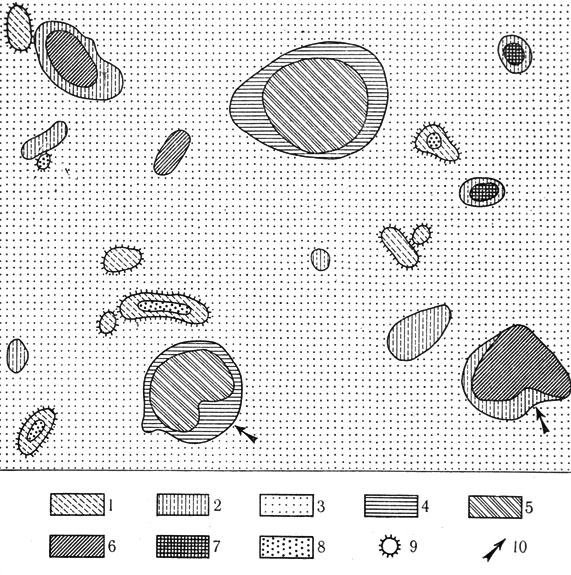
Рис. 22. Ассоциации пятнистого слабоостепненно-пустынного микрокомплекса (по Левиной, 1964): 1 - солянково-чернополынная на сусликовинах; 2 - типчаково-ромашниково белополынная; 3 - острецово-чернополынно-кокпековая на лугово-степных солончаковых солонцах наноповышенных участков; 4 - злаково-прутняково-ромашниковая по краям неглубоких микрозападин и в небольших нанозападинах на светло-каштановых солонцеватых почвах; 5 - типчаково-ковылковая с участием житняка; 6 - житняково-типчаково-тырсиковая; 7 - грудницево-типчаково-житняковая; 8 - острецовая с участием ромашника на сусликовинах; 9 - сусликовина; 10 - асимметричность топографии микрогруппировок растительности на месте руинных сусликовин
Выявление "руинных сусликовин" позволяет понять объективный ход развития сукцессии растительного покрова, имеющей в своей основе землеройную деятельность малых сусликов, и оценить общие размеры трансформирующего воздействия сусликов на растительность полупустыни.
В условиях засушливых зон с типичной для них разреженностью растительного покрова и относительно небольшой наземной фитомассой животные (как позвоночные, так и беспозвоночные) оказывают весьма существенное влияние на растительность, на объем фитопродукции, возобновление и распространение ряда видов, в том числе эдификаторов некоторых зональных сообществ (в полупустыне это черная полынь - Artemisia pauciflora). Расселение специфических комплексов зоогенной растительности связано с точками землеройной деятельности грызунов, нарушающих целостность первичного почвенно-растительного покрова.
В конкретных случаях воздействие животных оказывается различным: от прямого выедания растительной массы и повреждения семян и корневых систем до изменения среды жизни, включая "скотосбой" - вытаптывание растений и уплотнение почвы (преимущественно копытными), изменения микрорельефа и почвенно-гидрологических условий. Это касается в первую очередь грызунов и некоторых массовых видов беспозвоночных (таких, например, как в пустынях землероющие мокрицы), которые выступают как мощный биогеоценотический фактор и могут рассматриваться в качестве зонального фактора средообразования. Участие грызунов-землероев в формировании зональных биокомплексов и в поддержании процессов их биогеоценотического циклогенеза ярко проявляется в условиях полупустыни Северного Прикаспия, где малым сусликам принадлежит основная ценозо-регуляторная роль.
Существование зоогенных сукцессии растительности, связанное с деятельностью малых сусликов, специально отмечалось многими исследователями (Формозов, Ходашева, Голов, 1954; Ходашева, Динесман, 1961; Левина, 1961, 1964; Молчанова, 1973). Однако до настоящего времени анализ стадий сукцессионного процесса и выявление долголетних "следовых" изменений растительности, что важно для понимания механизма биогеоценотического процесса в полупустынях, не были проведены.
В обширной монографии по растительности полупустыни Северного Прикаспия Ф. Я. Левина (1964) посвящает специальный раздел зоогенным сменам, рассматривая их как род экзодинамических сукцессии. Ею отмечается ряд видов галофитного покрова молодых сусликовин (Lepidium perfoliatum, Salsola foliosa, S. brachiata), последующее преобладание разреженных группировок полукустарничков и развитие на заброшенных сусликовинах группировок ромашника со злаками. Однако картины зоогенной динамики смен растительности и причинных основ изменений структуры фитокомплексов в работе не дается. Особое внимание Ф. Я. Левина уделяет рассмотрению влияния на растительность степной пеструшки. Оно оказывается действительно существенным в годы вспышек массового размножения этого грызуна.
Однако глубина, длительность и постоянство влияния пеструшек несоизмеримы с масштабом средообразующей деятельности в полупустыне малых сусликов. И если изменения в растительном покрове, возникающие в годы высокой численности степной пеструшки, при всей их значительности и своеобразии нивелируются на протяжении нескольких лет и, таким образом, оказываются в масштабе процессов средообразования все же эпизодическими (они не вызывают глубоких перестроек почвенно-гидрологических условий и обширного ряда сопутствующих процессов), то землеройная деятельность малого суслика, охватывая практически всю продукционно-активную толщу зонального биогеоценоза полупустыни, выступает как фактор средообразования геологического масштаба, способный формировать облик территории и зонально обусловленный тип - схему узловых биогенных процессов в ландшафте полупустыни на протяжении тысячелетий.
Не анализируя все стороны ценозообразующей и геологической роли малых сусликов в полупустыне Прикаспия и оставляя в стороне вопросы потребления сусликами фитомассы, рассмотрим лишь то воздействие на растительность их роющей деятельности, которое проявляется в изменении состава и структуры растительного покрова и в последовательных этапах смены (сукцессии) растительности на бутанах сусликов и прилежащих к ним участках. Этапы - стадии зоогенной сукцессии растительности - суть результат сложного комплекса взаимосвязанных трансформацией рельефа, гидрологических условий и почв.
При высокой численности сусликовин, типичной для Прикаспия (местами до 200 и более на 1 га, включая нежилые и "руинные"), собственно зоогенные фитогруппировки на жилых бутанах и соседняя растительность, измененная также в результате деятельности малых сусликов, занимает большую часть территории, определяя комплексный характер растительного покрова и весь облик ландшафта прикаспийской полупустыни (см. рис. 23).
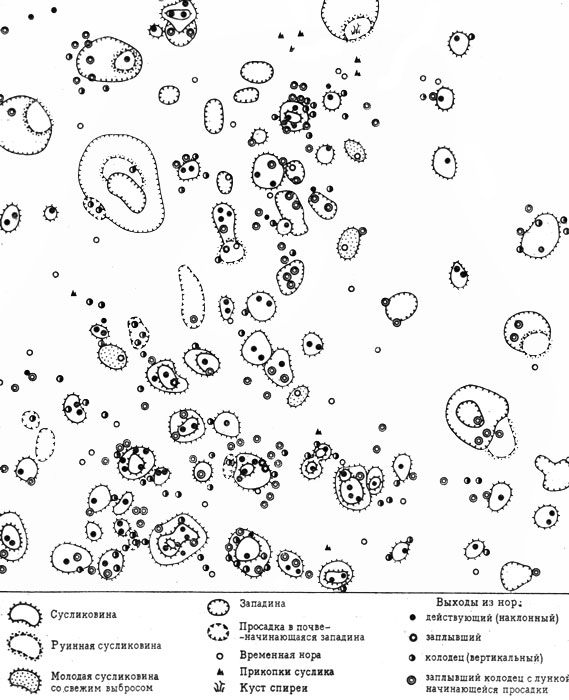
Рис. 23. Размещение сусликовин и выходов из нор малого суслика в глинистой полупустыне (Северный Прикаспий. Пробная площадь 0,25 га)
Установленный сукцессионный ряд включает 10 стадий, каждая из которых отличается своей спецификой (особенностями микрораспределения растений и структуры растительных группировок) и разной индивидуальной длительностью существования. В названии каждой стадии отмечается лишь одна из главных особенностей ее растительного покрова, что само по себе помимо приводимых ниже характеристик стадий облегчает выделение их на местности уже по физиономическим признакам. Стадии подразделяются на восходящий ряд развития сукцессии (I-IV стадии - обитаемые бутаны), на этап регенерирования первичного растительного покрова и возврата на сусликовину группировок коренной растительности солончаковых солонцов - ассоциаций черной полыни (V, или одна из более ранних стадий, если она представляет собой заброшенный необитаемый бутан, хорошо выделяемый в рельефе среди участков солончаковых солонцов) и на регрессивный ряд, когда на деградирующей, необитаемой сусликовине, подверженной эрозионным процессам и глубоким изменениям почвенного покрова, или уже на осевшем остаточном бутане развиваются группировки растений сухостепного и даже лугово-степного характера (VI-X стадии - заброшенные сусликами, давно необитаемые и "руинные сусликовины" уже утратившие форму бутана и иногда лишь слабо выделяющиеся в рельефе на стадиях VIII-X).
- I стадия "свежего выброса почвы" на первичной, неизмененной растительности солончакового солонца.
- II стадия "бордюрного кольца растений" вокруг молодой сусликовины.
- III стадия "разнотравного бугра" средневозрастной сформировавшейся сусликовины над обитаемой норой.
- IV стадия "солянковых маяков" (перезревший бутан с функционирующей норой, расположенной асимметрично лишь на части бугра).
- V стадия "полынного поглощения" (заброшенный необитаемый сусликами бутан, на который возвратились группировки коренной чернополынной растительности с окружающих солончаковых солонцов).
- VI стадия "злаково-ромашниковой подковы" (отмирающий необитаемый бутан асимметрично с двух (или с одной) сторон окружен измененной растительностью, поселяющейся в понижениях - просадках, возникших на месте старых вертикальных ходов вокруг сусликовины. Группировки растительности резко асимметричны. Одной из сторон старый бутан смыкается с прутняково-полынной коренной растительностью соседнего солончакового солонца).
- VII стадия "злакового опоясывания" (необитаемый, слегка осевший бутан окружен почти полностью или полностью, кольцом злаков, растущих в просадках почвы, сливающихся в серпообразную западину. При этом сохраняется асимметрия растительного покрова).
- VIII стадия "центрирования бордюрного комплекса растительности западин на руинном бутане" (сильно осевший, иногда слабо заметный из-за окружающих высоких злаков уплощенный бугор бывшего бутана, зарастающий в центре своеобразной группировкой видов, типичных для краевой зоны больших долговременно существующих западин и падинных понижений лиманного типа: майский полынок (Artemisia austriaca), грудница волосистая (Linosyris villosa), молочай волнистый (Euphorbia undulata), оносма (Onosma setosum) единично, прутняк (Kochia prostrata), острец (Eneurolepidium ramosum), татарник (Onopordum acanthium), мятлик луковичный (Роа bulbosa), тонконог (Koeleria glauca), астрагалы (Astragalus brachylobus et A. virgatus), кермек сарептский и изредка тысячелистник (Achilea nobilis), костер (Bromus tectorum) и др.).
- IX стадия "злакового поглощения" (заметный в рельефе, но уплощенный и осевший руинный бутан, зарастающий почти сплошь злаками, главным образом житняком пустынным (Agropyron desertorum) и острецом. При этом все же отмечается концентрическое расположение растительности на бывшем бутане и асимметрия ее группировок прежнего типа. В окружающей западине в разных соотношениях господствуют житняк гребневидный (Agropyron pectiniforme) и житняк пустынный.
- X стадия "кустарниковых куртин" (совершенно осевший бутан, скрытый злаками: житняками и ковылями и одним-двумя кустами или даже зарослями таволги зверобоелистной (Spirea hypericifolia), обнаруживающийся под покровом этой растительности на глаз или при нивелирной съемке внутри округлой или слегка вытянутой западины, но всегда несколько ближе к ее краю, что повторяет специфическую асимметрию растительных группировок, связанную с генезисом бутана и особенно четко выраженную на стадиях VIII и IX).
Ход сукцессии определяется до IV стадии роющей деятельностью сусликов. Следует однако учитывать, что суслики нередко забрасывают свои бутаны на любой стадии, и тогда начинаются процессы регенерации почв и возврата коренной растительности солончаковых солонцов (до V и даже VI). В годы высокой численности, вновь поселяясь на заброшенной ранее и "постаревшей" сусликовине (если даже это была молодая или средневозрастная), суслики способны оживить и активизировать замерший циклогенез, остановив происходящую регенерацию чернополынной растительности окружающих солонцов и стимулируя дальнейшие этапы сукцессии. Однако со стадии VI и далее смены растительных группировок происходят автономно, без непосредственного участия сусликов, целиком на основе геоморфологических и почвенно-гидрологических процессов, обусловленных тем не менее предыдущей роющей деятельностью зверьков, предопределившей полностью направление, характер и этапы не только смен растительности, но и закономерных трансформаций всего биогеоценотического комплекса, типичного для прикаспийской полупустыни.
После четырех стадий, когда суслики оставляют развитый бутан, процесс разделяется в зависимости от начального положения сусликовины в мезорельефе, размера водосборной площади и возможностей развития просадок поверхности. В первом случае при расположении сусликовины на наиболее повышенных участках солончаковых солонцов, где увлажнение атмосферными осадками минимально (в силу сдувания снегового покрова, большего промерзания почвы и лучшего стока) и где, следовательно, промывания почвы не происходит, начинается относительно быстрая регенерация первичных чернополынных группировок.
Во втором случае, если сусликовина находится на более пониженных склоновых участках солончаковых солонцов, в полосе, соседствующей с падинами, где несколько лучше условия увлажнения почвы, наблюдается обычное образование просадок поверхности в зоне разброса вертикальных нор. Здесь сукцессия растительности получает дальнейшее автономное развитие, охватывая площадь не только остаточной оседающей сусликовины, но и прилегающие участки с выходами полузасыпанных вертикальных нор. Эта ветвь сукцессии характеризуется тенденцией к спонтанному расширению своей площади, однако в пределах, определяемых моментом остановки развития просадок и сформированием относительно плотного злакового задернения.
При изменении площади, охваченной сукцессией, общая форма ее и конфигурация границ растительных группировок внутри сукцессионного поля сохраняет свою первичную зоогенную схему асимметрии. Эта ветвь сукцессии характеризуется развитием и сменой группировок злаков и светло-каштановых почв, поглощающих в итоге руинный бугорок бывшей сусликовины. Нередко именно на нем поселяются куст или целая заросль спиреи зверобоелистной. В таких зарослях обычно возникают колонии обыкновенных полевок и устраивают норы домовые мыши.
Анализ стадий зоогенной сукцессии растительности глинистой полупустыни (Северный Прикаспий)
Стадия I - свежего выброса почвы на первичной, неизмененной растительности. Возраст 1-7-10 лет. Показатель возраста - наличие вертикальных ходов-"колодцев" вокруг выброса, иногда похожего внешне на временную нору, но все же представляющего собой начальную стадию сусликовины, поскольку вертикальный ход указывает на присутствие гнездовой камеры, из которой суслик выходит весной, после зимней спячки, прорывая каждый раз новый "колодец". Поэтому число "колодцев" или их остатков равно количеству зим и на год или два меньше возраста сусликовины, поскольку в первый год сусликовина могла использоваться лишь как временная нора.
Почвенный выброс ежегодно обновляется и почти не зарастает растительностью. Растения, свойственные нано- и микроповышениям с солончаковыми солонцами, преимущественно кусты черной полыни, отчасти прутняка, прорастают выброшенную толщу почвы. Бордюрное кольцо растений практически не выражено. Проективное покрытие на выбросе 5-10%. Мощность развития куста полыни несколько превышает таковую на участке соседнего солончакового солонца в связи с лучшим влагонакоплением в весеннее время на выбросе рыхлой почвы с еще не сформированной структурой. Свежий выброс, как правило, обогащен легкорастворимыми солями. "Сумма солей в нем достигает 1,68%. Преобладают ионы SO4" (19,9 мг*экв/100 г почвы), Na· (14,95), Са¨ (6,47) и Cl' (4,99). Характерно, что ионов Na· содержится здесь в 2 раза больше, чем Са¨. Особенно много выносится на поверхность гипса. В свежевыброшенном материале количество его, оцениваемое по содержанию нерастворимого в воде SO4", достигает 23 мг*экв/100 г почвы, а под выбросом в надсолонцовом и солонцовом горизонтах не превышает 3,2 мг*экв (Зубкова, 1971)*. Соленасыщенность свежих выбросов, очевидно, мешает их быстрому зарастанию.
* (Изучение химического состава почв "молодых", "средневозрастных" и "старых" сусликовин, заброшенных сусликами, и влияния выбросов на химические свойства солончаковых солонцов было проведено Л. В. Зубковой на Джаныбекском стационаре АН СССР (Зубкова, 1971). Это дает возможность идентифицировать II, III и IV стадии сукцессионного ряда с изменениями солевого режима почв. Исследований почв V-X стадий не проводилось.)
Стадия II - бордюрного кольца растений вокруг молодой сусликовины (см. рис. 24). Возраст 10-20 лет. Выбросы почвы за ряд лет образовывают относительно небольшой по размерам бугорок диаметром 1,0-1,5 м и высотой 10-15 см. Его особенность в отличие от "первичного выброса" состоит в том, что не вся поверхность бугра покрыта свежеперерытой почвой, поднятой в текущем году. Следовательно, в периферийных частях молодого бутана, там, где суслики не нарушили толщу ранее вынесенной ими почвы вторичным ее перекапыванием, начинается специфический почвообразовательный процесс в "перемешанной" зоогенной почвенной массе над погребенной поверхностью солончакового солонца.

Рис. 24. Растительность сусликовин малого суслика на стадии 2 ('бордюрного кольца растений')
На молодом бутане поселяются растения, корневая система которых развивается уже в толще вынесенной сусликами почвы и вместе с тем проникает под погребенную прежнюю поверхность надсолонцового горизонта. Во влажные годы на молодом бутане хорошо приживается семенное возобновление черной полыни и прутняка.
Состав видов: черная полынь (эдификатор), прутняк и мятлик луковичный (реже). Кусты черной полыни достигают мощного развития и большей высоты, чем на соседнем солонце, не затронутом деятельностью землероев, что объясняется лучшей влагообеспеченностыо молодого бутана. Более высоким оказывается и содержание влаги в каждом кусте полыни на молодом бутане по сравнению со сравнимым кустом ее на солончаковом солонце (влажность средней пробы по 40 стеблей годичного прироста черной полыни, собранных с молодой сусликовины в августе, равнялась 69%, а пробы с соседнего участка солончакового солонца - 57%).
Для молодого бутана характерно относительно хорошее развитие эфемеров, во влажные годы - в особенности дескурайнии (Descurainia sophia) и отчасти перечника пронзенно-листного (Lepidium perfoliatum), в сухие - мортука пшеничного (Eremopyrum triticeum) и бурачка (Alissum desertorum). Достаточно обычен на молодых бутанах прутняк и в меньшей степени, по их периферии, - солянка многолистная.
Главная особенность растительного покрова молодого бутана - ясно выраженное "бордюрное кольцо", образованное особенно мощно развитыми кустами черной полыни и прутняка, часто с участием мятлика луковичного, растущими загущенными куртинками по его периферии, в зоне контакта "молодой почвы" и солончакового солонца. Иногда в бордюрном кольце растут мортук пшеничный и мортук восточный (Eremopyrum orientate), рогоплодник песчаный - эбелек (Ceratocarpus arenarius) и весной - гусиные луки (Gagea bulbifera et G. pusilla).
Средняя высота стеблей годичного прироста черной полыни (пробы по 40 стеблей с каждого курганчика взяты в августе) на вершине молодой сусликовины составила 19,2 см, а в ее бордюрном кольце - 23,7 см, при максимуме - 29,0 см. Проективное покрытие на молодой сусликовине-15 - 20% (иногда 25%), в бордюрном кольце - 50 - 70%. Заселение ее собственной растительностью в особенности по периферии и на склонах коррелируется с началом почвообразовательного процесса в толще вынесенного сусликами на дневную поверхность почвенного материала, утратившего свою первичную структуру и насыщенного солями.
Главная особенность почвообразования в вынесенном зоогенном материале - начало вымывания солей атмосферными осадками и происходящее параллельно с этим быстрое изменение соотношения различных ионов в молодой почве формирующейся сусликовины. Содержание гипса на этой стадии в молодых сусликовинах существенно сокращается, по данным Л. В. Зубковой (1971), до 8,2 мг экв/100 г почвы. Сумма солей в молодой сусликовине уменьшается до 0,5%, т. е. втрое по сравнению со свежим выбросом. Одновременно в надсолонцовом горизонте погребенной почвы общее содержание солей возрастает с 0,08 до 0,52%.
В связи с тем что скорость выноса солей натрия оказалась выше, чем солей кальция, соотношение их быстро изменяется. И если в свежем выбросе (I стадия) содержание солей натрия вдвое превышало количество солей кальция, то в почве молодой сусликовины, где уже происходят процессы промывания и сноса атмосферными осадками, напротив, содержание кальция становится в 2 раза больше, чем натрия, что свидетельствует о мелиоративном эффекте (Ходашева и Динесман, 1961; Зубкова, 1971), начинающемся на II стадии биогеоценотических трансформаций "сусликового комплекса". При этом мелиорируется не только молодая почва, но отчасти и погребенный под ней солонцовый горизонт, где также происходит некоторое накопление солей кальция.
Хорошее развитие бордюрного кольца растений, образованного главным образом теми же видами, которые населяют соседние участки солончаковых солонцов, объясняется эффектом частичной мелиорации узкой концентрической полоски солонца, примыкающей к подножию сусликовины, и, вероятно, несколько лучшей влагообеспеченностью периферийной части сусликовины. Происходящее в контактной зоне молодой сусликовины и солонца слабое рассоление достаточно благоприятно для черной полыни, прутняка, мятлика луковичного и некоторых эфемеров по сравнению с условиями соседнего солонца, что обусловливает их загущенное произрастание в "бордюрном кольце" и пышное развитие отдельных кустов. Однако этой узколокальной и относительно слабой мелиорации оказывается мало, чтобы обеспечить существенное или коренное изменение состава видов растений как в краевых участках, так и на поверхности начавшей рассоляться молодой сусликовины.
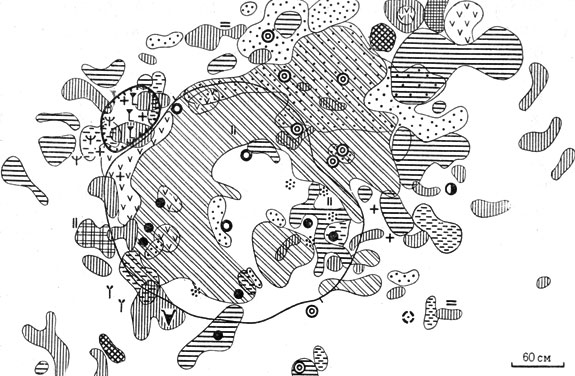
Рис. 25а. Растительность сусликовин малого суслика на стадии 3 ('разнотравного бугра') (условные обозначения, как на рис. 24)
Стадия III - разнотравного бугра - средневозрастная сформировавшаяся (зрелая) сусликовина над обитаемой норой (см. рис. 25). Возраст от 20 и более до нескольких десятков лет.
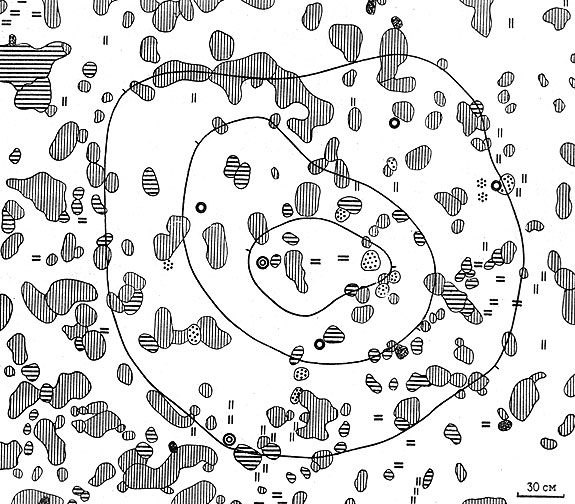
Рис. 25б. Растительность сусликовин малого суслика на стадии 5 ('полынного поглощения') (условные обозначения, как на рис. 24)
Зрелая сусликовина достигает средних и максимальных размеров, в полупустыне Волжско-Уральского междуречья диаметр ее 2-4 м при высоте 20-40 см (Ф. Я. Левина (1964) указывает максимум высоты 50 см). Сусликовина резко выделяется среди растительности солончакового солонца (ассоциации черной полыни, преимущественно прутняково-чернополынные, камфоросмово-чернополынные и др.) разнообразным набором видов растений, хорошо развитым и загущенным травостоем. Свежие же прикопки сусликов остаются оголенными.
На поверхности сусликовины доминирует ксерофильное разнотравье, включая некоторые сорняки, весной и в самом начале лета вегетируют эфемеры (во влажные годы обильно). В большинстве случаев несколько видов образуют более или менее плотные куртины, делящие поверхность бугра на отдельные сегменты: главным образом перечник пронзеннолистный (Lepidium perfoliatum), липучка прямая (Lappula stricta), эфемер дескурайния (Descurainia sophiae). Иногда один из этих видов мощно разрастается, захватывая практически всю сусликовину, и лишь ее периферия выделяется разнообразием трав, образующих прерывистое бордюрное кольцо. Среди видов, сопутствующих названным доминантам, на вершине и склонах зрелой сусликовины наблюдаются субдоминанты: острец (Eneurolepidium ramosum), мятлик луковичный (Роа bulbosa), ромашник (Pyrethrum achilleifolium), прутняк (Kochia prostrata). Отдельными особями на стадии "разнотравного бугра" встречаются сорняки: марь белая (Chenopodium album), лебеда татарская (Atriplex tatarica) и редко (A. sphaeromorpha) яснотка стеблеобъемлющая (Lamium amplexicaule), кермек Гмелина (Limonium gmelinii), горец отклоненный (Polygonum patulum), мортук пшеничный (Eremopyrum triticeum), рогоглавник пряморогий (Ceratocephalus orthoceras), реже бурачок (Alissum desertorum), шизмус (Schistnus arabicus), тонконог (Koeleria gracilis) и тырсик (Stipa sareptana), ранневесенний астрагал (Astragalus contortuplicatus), эфемер однолетник и ферула голая (Ferula nuda). Изредка можно обнаружить отдельные кустики черной полыни, не образующие здесь многолетних куртин.
Бордюрное кольцо обычно относительно узкое, в нем преобладают ромашник, рогоплодник песчаный - эбелек, очень типичны липучка прямая, мортук восточный и мортук пшеничный (в бордюре обильнее, чем на вершине сусликовины), прутняк, черная полынь, мятлик луковичный, рогоглавник пряморогий, изредка костер и отдельные растения эхинопсилона очитковидного (Echynopsilon sedoides) и однолетника крупки (Draba nemorosa). В случае близости к бордюру просадки в почве вокруг заплывшего вертикального хода нередко добавляются вероника (Veronica verna), желтушник (Erysimum leocanthemum) и житняк пустынный (Agropyron desertorum). Набор видов бордюрного кольца характеризуется большим разнообразием (10-15 видов) и мало отличается от состава растительного покрова поверхности самой сусликовины на стадии разнотравного бугра (12-17 видов, из них 10-12 общие с бордюрным кольцом и лишь 2-3 вида общие с растительным покровом солончакового солонца). Здесь сочетаются виды - ксеромезофилы и такие широко распространенные в аридных зонах ксерофилы, как эбелек, мортук восточный и рогоглавник.
Однако высота и мощность развития растений и степень загущенности их в бордюрном кольце на стадии разнотравного бугра мало отличается от тех же показателей на основной поверхности сусликовины. Кроме того, кольцо прерывисто и асимметрично, наиболее выражено оно с той стороны бутана, где суслики омолаживают его поверхность, вынося из глубины новую почву. Таким образом, кольцо растений на стадии III характеризуется значительным качественным разнообразием состава видов, но почти не выделяется обилием и высотой растений, чем принципиально отличается от стадии II (бордюр которой включает всего 2-3 вида, тех же, что на соседнем солонце). Обилием же особей и габитусом растений бордюрное кольцо на стадии II выделяется значительно сильнее. Проективное покрытие: на поверхности сусликовины - 50-75%, в бордюрном кольце (прерывистом, выраженном не всегда четко) - 60-85%.
Особенности состава растительного покрова зрелой сусликовины на стадии разнотравного бугра, включающего эвримезофитные, ксеромезофитные, галомезофитные и эвригалофитные виды, - это повышенные показатели обилия и хорошее развитие индивидов по сравнению с растениями на ненарушенных солонцовых почвах и на заброшенных старых сусликовинах. Такой характер покрова можно объяснить дальнейшими специфическими зоогенными трансформациями в почве бутана, связанными с изменением водного и солевого режима, с процессами вымывания и миграции солей, с изменением соотношения ионов кальция и натрия в толще выброшенной сусликами почвы, а также и в погребенном солонцовом горизонте в пользу кальция. Процессы рассоления и дальнейшей естественной мелиорации поверхностных слоев типичны для данной стадии. В разные по увлажнению годы темпы этого процесса, очевидно, существенно отличаются.
Почвенный профиль сформировавшейся жилой сусликовины специфичен и резко отличается от такового как на ненарушенном сусликами солончаковом солонце, так и на молодом бутане. Химические свойства солончакового солонца, оказавшегося погребенным под развитым бутаном, также существенно изменяются. Соли из выброшенного сусликами почвенного материала атмосферной влагой частично вымываются и накапливаются в погребенных надсолонцовом и солонцовом горизонтах. В зрелых обитаемых сусликовинах "содержание солей в верхнем слое (0-9 см) уменьшается по сравнению с молодой сусликовиной в 3,5 раза. В нижележащих слоях сусликовины (и в выбросе, и в сомкнувшемся с ним погребенном надсолонцовом горизонте) содержание солей по сравнению с молодой сусликовиной несколько увеличивается (с 0,56 до 0,76-0,91%). Возрастает оно и в погребенном солонцовом горизонте по сравнению с ненарушенной почвой (с 1,01 до 1,70%)" (Зубкова, 1971). В надсолонцовом и солонцовом горизонтах накапливаются помимо сульфатов и хлориды.
В толще выброшенной почвы в сусликовине отношение натрия к кальцию последовательно уменьшается, снижаясь до 0,3.
В погребенном солонцовом горизонте содержание кальция увеличивается по сравнению с ненарушенной почвой с 0,4-0,7 до 5,8 мг*экв/100 г почвы, тогда как содержание натрия почти не изменяется. Таким образом, отношение натрия к кальцию в надсолонцовом и солонцовом горизонтах уменьшается с 18,6 до 2,4. Содержание гипса (определяется по количеству нерастворимого в воде SO4") в поверхностном слое почвы - 0,9 см. На средневозрастной сусликовине оно уменьшается до 1,6 мг*экв/100 г почвы по сравнению с 23 мг*экв/100 г почвы в свежевыброшенном на поверхность материале.
Следовательно, выявленные и описанные выше изменения растительности зрелой ("средневозрастной") сусликовины могут быть объяснены рассолением приповерхностного слоя молодой почвы бутана, выносимой сусликами на протяжении десятков лет, лучшей ее влагообеспеченностью в периоды увлажнения атмосферными осадками и благоприятными для растительности изменениями солевого режима погребенной под бутаном почвы. Зоогенная мелиорация солончакового солонца, получающая заметное развитие впервые на этой стадии биогеоценотического цикла, основывается прежде всего на локальном изменении водного режима в результате весьма значительной землеройной деятельности малых сусликов, формирующей своеобразный мелкобугорчатый "сусликовый рельеф" полупустыни.
Рельефообразующая деятельность сусликов в полупустыне способствует организации стока, перераспределению атмосферной влаги и изменению влагообеспеченности отдельных участков. Условия промачивания и промывания почв сказываются на перемещении почвенной влаги, явлениях капиллярности почв и на различном испарении с поверхностей, измененных землероями.
Глубокие изменения растительности (ее состава и обилия) на стадии "разнотравного бугра" оказываются результатом и одновременно индикатором сложного комплекса почвенно-гидрологических процессов, в основе которых лежит длящаяся десятилетия землеройная деятельность сусликов (явление "зоогидропедоморфоза"). В итоге разнообразие растительного покрова сусликовины на стадии III в 5-7 раз превосходит состав видов на солонце и на сусликовине II стадии за счет главным образом ксеромезофильных форм и эфемеров. Показатели обилия в 3-4 раза превышают таковые на солонце и в покрове на стадии II.
Длительность течения стадии разнотравного бугра, равная иногда многим десяткам лет, ограничивается моментом, когда суслики надолго покинут "перезревшую" сусликовину и процессы изменения почвы и растительности здесь пойдут в ином направлении. В процессе же развития крупной сусликовины в пределах стадии III происходит неравномерное старение одних периферийных участков и "омоложение" других - тех, где суслики продолжают активно перекапывать и выносить на поверхность почву. В результате возникает типичная для стадии разнотравного бугра асимметрия растительного покрова и наблюдается изменчивость вариантов разнотравья с довольно характерным увеличением в травостое мари, лебеды, татарника, молочая волнистого. Одновременно появляются однолетники - солянка многолистная и петросимония однотычинковая.
Стадия IV - солянковых маяков. Возраст около 40-50 лет и более. Перезревший бутан с периодически функционирующей асимметрично расположенной на его склоне норой. Происходит дальнейшее рассоление выброшенного на поверхность материала с уменьшением содержания солей до 0,05%, при одновременном накоплении солей в погребенном солонцовом горизонте и в нижней части почвенного профиля под перезревшей сусликовиной, начиная с верхнего подсолонцового горизонта. Последнее может происходить за счет подтягивания солей снизу в связи с особенностями водного режима таких сусликовин. Появление на бутане на этой стадии крупных, издали заметных солянок - лиственничной и итцегека (S. aphilla), имеющих глубокую корневую систему, свидетельствует об увеличении соленосности солонцового и подсолонцового горизонтов.
Стадия V - "полынного поглощения". Представляет собой возвращение чернополынной растительности на поверхность заброшенного бутана и регенерацию ее первичных группировок, что соответствует процессу восстановления в толще давно вынесенной почвы солевого режима, близкого к таковому на ненарушенном солончаковом солонце. Важное значение для коренного чернополынника имеет сходство солевого состава верхних горизонтов почвы над солонцовым горизонтом (0-20 см), в которых непосредственно сосредоточивается основная масса корней черной полыни. Сравнение содержания обменных оснований в почве в надсолонцовом горизонте ненарушенного солончакового солонца и в почве заброшенной сусликовины (в горизонте 0-10 см) указывает на их экологическую равноценность (соответственно: Са ¨ - 11,6 и 13,2 мг*экв/100 г почвы; Mg ¨ - 3,2 и 2,8; К · - 2,05 и 1,61; Na · - 1,44 и 1,44; сумма солей - 18,29 и 19,05 мг*экв/100 г почвы (Зубкова, 1971). Этой стадией завершается первый короткий разнотравный ряд сукцессионных преобразований растительности.
"Злаковый ряд" сукцессии от стадии "солянковых маяков" ведет непосредственно к стадии "злаково-ромашниковой подковы" (VI, см. рис. 26) и связан в своем развитии с постепенным переходом от непромывного водного режима почв к периодически промывному на стадиях "злакового поглощения" (IX) и "кустарниковых куртин" (X), когда появляется и наращивается гумусовый горизонт. Этапы смены растительности соответствуют постепенному формированию в растущих просадках светло-каштановых солонцеватых почв на стадиях: "злаково-ромашниковой подковы" (VI), "злакового опоясывания" (VII) и "центрирования бордюрного комплекса растительности западин на руинном бутане" (VIII) и в итоге темноцветных черноземовидных почв в западинах с покровом из злаков (житняков и ковылей) и куртинами кустарников: таволги зверобоелистной и городчатой (IX и X).
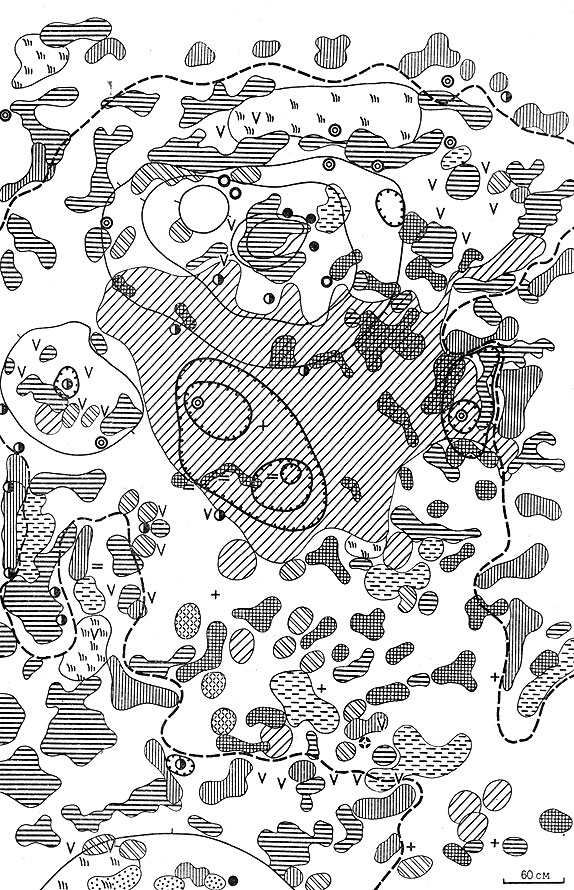
Рис. 26. Растительность сусликовин малого суслика на стадии 6 ('злаково-ромашниковой подковы') (условные обозначения, как на рис. 24)
Итак, зоогенная сукцессия растительности в глинистой полупустыне имеет основой сусликовины, представляющие собой точки нарушения плоскостного рельефа, почвенно-гидрологических условий и целостности первичного растительного покрова. Основная особенность роющей деятельности малых сусликов в отличие от других видов грызунов в этой природной зоне - охват всей толщи биогеоценоза (всех его горизонтов), что позволяет рассматривать землеройную деятельность этого вида в условиях семиаридного климатического режима с атмосферным увлажнением, допускающим частичное промывание почв, как важнейший зональный фактор средообразования.
Деятельность сусликов служит стартовым моментом для почвенно-гидрологических процессов, которые затем развиваются автономно и отражаются в структуре биогеоценоза и в специфической топографии растительных группировок. В этом смысле малые суслики "сделали" прикаспийскую полупустыню. Типичная для Северного Прикаспия, легко выделяемая подковообразная асимметрия концентрической топографии растительных группировок представляет собой "сусликовое клеймо", сохраняющееся в схеме структурной организации биогеоценотического покрова на разных стадиях его развития.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animal.geoman.ru/ 'Мир животных'